Digestive systems
Digestive systems take many, many forms. There is a fundamental distinction between internal and external digestion. External digestion was the first to evolve, and most fungi still rely on it. In this process, enzymes are secreted into the environment surrounding the organism, where they break down an organic material, and some of the products diffuse back to the organism. Later, animals evolved by rolling into a tube and acquiring internal digestion, which is more efficient because more of the broken down products can be captured, and the chemical environment can be more efficiently controlled.
Some organisms, including nearly all spiders, simply secrete biotoxins and digestive chemicals (e.g., enzymes) into the extracellular environment prior to ingestion of the consequent "soup". In others, once potential nutrients or food is inside the organism, digestion can be conducted to a vesicle or a sac-like structure, through a tube, or through several specialized organs aimed at making the absorption of nutrients more efficient.
Some organisms, including nearly all spiders, simply secrete biotoxins and digestive chemicals (e.g., enzymes) into the extracellular environment prior to ingestion of the consequent "soup". In others, once potential nutrients or food is inside the organism, digestion can be conducted to a vesicle or a sac-like structure, through a tube, or through several specialized organs aimed at making the absorption of nutrients more efficient.
Secretion systems
Bacteria use several systems to obtain nutrients from other organisms in the environments.
Channel transport system
In a channel transport system several proteins form a contiguous channel traversing the inner and outer membranes of the bacteria. It is a simple system, which consists of only three protein subunits: the ABC protein, membrane fusion protein (MFP), and outer membrane protein (OMP)[specify]. This secretion system transports various molecules, from ions, drugs, to proteins of various sizes (20 - 900 kDa). The molecules secreted vary in size from the small Escherichia coli peptide colicin V, (10 kDa) to the Pseudomonas fluorescens cell adhesion protein LapA of 900 kDa.
Molecular syringe
One molecular syringe is used through which a bacterium (e.g. certain types of Salmonella,Shigella, Yersinia) can inject proteins into eukaryotic cells. One such mechanism was first discovered in Y. pestis and showed that toxins could be injected directly from the bacterial cytoplasm into the cytoplasm of its host's cells rather than simply be secreted into the extracellular medium.
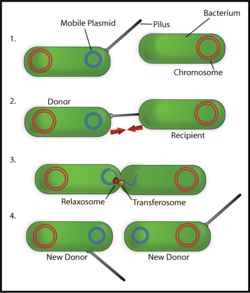
Conjugation machinery
The conjugation machinery of some bacteria (and archaeal flagella) is capable of transporting both DNA and proteins. It was discovered in Agrobacterium tumefaciens, which uses this system to introduce the Ti plasmid and proteins into the host which develops the crown gall (tumor).[6] The VirB complex of Agrobacterium tumefaciens is the prototypic system.
The nitrogen fixing Rhizobia are an interesting case, wherein conjugative elements naturally engage in inter-kingdom conjugation. Such elements as the Agrobacterium Ti or Ri plasmids contain elements that can transfer to plant cells. Transferred genes enter the plant cell nucleus and effectively transform the plant cells into factories for the production of opines, which the bacteria use as carbon and energy sources. Infected plant cells form crown gall or root tumors. The Ti and Ri plasmids are thus endosymbionts of the bacteria, which are in turn endosymbionts (or parasites) of the infected plant.
The Ti and Ri plasmids are themselves conjugative. Ti and Ri transfer between bacteria uses an independent system (the tra, or transfer, operon) from that for inter-kingdom transfer (the vir, or virulence, operon). Such transfer creates virulent strains from previously avirulentAgrobacteria.
The nitrogen fixing Rhizobia are an interesting case, wherein conjugative elements naturally engage in inter-kingdom conjugation. Such elements as the Agrobacterium Ti or Ri plasmids contain elements that can transfer to plant cells. Transferred genes enter the plant cell nucleus and effectively transform the plant cells into factories for the production of opines, which the bacteria use as carbon and energy sources. Infected plant cells form crown gall or root tumors. The Ti and Ri plasmids are thus endosymbionts of the bacteria, which are in turn endosymbionts (or parasites) of the infected plant.
The Ti and Ri plasmids are themselves conjugative. Ti and Ri transfer between bacteria uses an independent system (the tra, or transfer, operon) from that for inter-kingdom transfer (the vir, or virulence, operon). Such transfer creates virulent strains from previously avirulentAgrobacteria.
Release of outer membrane vesicles
In addition to the use of the multiprotein complexes listed above, Gram-negative bacteria possess another method for release of material: the formation of outer membrane vesicles. Portions of the outer membrane pinch off, forming spherical structures made of a lipid bilayer enclosing periplasmic materials. Vesicles from a number of bacterial species have been found to contain virulence factors, some have immunomodulatory effects, and some can directly adhere to and intoxicate host cells. While release of vesicles has been demonstrated as a general response to stress conditions, the process of loading cargo proteins seems to be selective.
Gastrovascular cavity
The gastrovascular cavity functions as a stomach in both digestion and the distribution of nutrients to all parts of the body. Extracellular digestion takes place within this central cavity which is lined with the gastrodermis, the internal layer of epithelium. This cavity has only one opening to the outside that functions as both a mouth and an anus: waste and undigested matter is excreted through the mouth/anus, which can be described as an incomplete gut.
In a plant such as the Venus Flytrap that can make its own food through photosynthesis, it does not eat and digest its prey for the traditional objectives of harvesting energy and carbon, but mines prey primarily for essential nutrients (nitrogen and phosphorus in particular) that are in short supply in its boggy, acidic habitat.
In a plant such as the Venus Flytrap that can make its own food through photosynthesis, it does not eat and digest its prey for the traditional objectives of harvesting energy and carbon, but mines prey primarily for essential nutrients (nitrogen and phosphorus in particular) that are in short supply in its boggy, acidic habitat.

